| Acerca de... |
| Geocities |
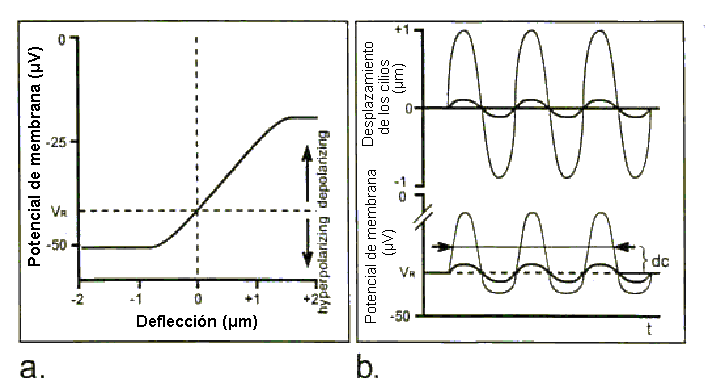 Fig. 26. A: Transducción característica función de entrada-salida. B: Potencial receptor intracelular para relativamente bajos versus altos desplazamientos ciliares. Toda esta fantástica bioingeniería creada por la madre naturaleza no solo sirve para poseer un mecanismo sensorial muy sensible, ¡sino también para producir un mecanismo que pueda responder selectivamente a sonidos de distintas frecuencias mientras representa mucho mas precisamente el contenido de frecuencias de sonidos más complejos!. La micromecánica es aun área de interés candente. Quedan otros misterios por resolver guardando los detalles de cómo exactamente la respuesta móvil de la CCE interactúa con la mecánica de la M.B. La contribución relativa de procesos activos versus procesos pasivos (sin mencionar el debate de cual proceso es activo versus uno pasivo). Otro tema intrigante es la posible regulación de este proceso por el sistema nervioso central (SNC). Resumen Una cantidad significativa de procesamiento de señal ocurre atreves de los mecanismos de la cóclea, antes de cualquier codificación neuronal. El espectro sonoro a sido, en efecto, transformado desde coordenadas "amplitud v/s frecuencia" a una "amplitud v/s distancia", con la distancia correspondiente al lugar máximo de la onda viajera. Así, por ejemplo, la respuesta de las células ciliadas situadas en un lugar específico a lo largo de la M.B. se parece a la respuesta de un filtro pasa-bandas dada una señal de ancho de banda a la entrada de la cóclea. La cadena total de los eventos macromecánicos conducen a los eventos micromecánicos que en realidad involucran mas que la cóclea, como se ilustra en la fig. 27. El sonido impreso sobre la membrana timpánica ajusta la cadena osicular a la vibración, la cual, en cambio, causa el desplazamiento de la M.B. e inicia la propagación de la onda viajera a lo largo de esta. Puede esperarse que, ya que la energía del sonido pasa a través de otros mecanismos antes de entrar a la cóclea, los mecanismos afectaran la eficiencia del acoplamiento de energía del sistema coclear. Consecuentemente, el espectro de sonido que llega a la cóclea se espera esté algo contaminado. Efectivamente, la influencia de las porciones externas y medias del oído sobre el espectro de sonido que alcanza a la cóclea es substancial. Esta parte del sistema, de hecho, determina que sonidos son audibles y cuan bien son escuchados. 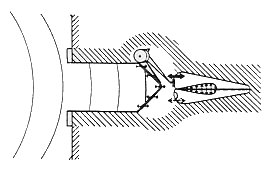 Fig. 27. Diagrama altamente esquematizado de todos los eventos conducidos desde que ingresa el sonido en el O.E. hasta la excitación de las ondas viajeras en la cóclea. Transducción A los órganos sensoriales le conciernen tres procesos:
En el sistema auditivo (sistema sensorial secundario), la señal receptora es generada en la célula ciliada y se denomina, "Corriente receptora". Ante los potenciales receptores generados por las células ciliadas, la neurona asume básicamente 1 de 2 estados: On u Off. Ni la altura ni la duración de los impulsos neuronales, lleva transformaciones más significantes una de la otra, ya que, sus dimensiones son bastante uniformes. 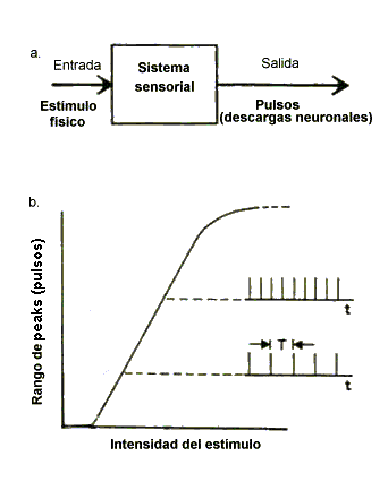 Fig. 16. A: Representación tipo caja negra de los sistemas sensoriales en general. B: Cuadro hipotético del rango de peaks (pulsos) como función de la intensidad del estímulo. Consecuentemente el primer aspecto de la información entregada por estos pulsos es cuan a menudo ellos ocurren, mejor denominado, su frecuencia o Rango de peaks (RP). (Estos pulsos tienen muy cortas duraciones alrededor de 1 mseg, así que, ellos lucen como peaks cuando son grabados y monitoreados con un osciloscopio. Como se muestra en la fig. 16 b, el RP aumenta a medida que aumenta la intensidad del estímulo, así la intensidad es codificada esencialmente como una frecuencia. Introducción a los modelos cocleares La utilización de modelos físicos y matemáticos, ha sido muy importante para establecer la existencia de ondas viajeras en la cóclea como un hecho, más que solo otra teoría. Aunque es posible hacer que un modelo haga lo que uno desea, los modelos, por muy cuidadosamente construidos, pueden tener problemas obvios de interpretación de datos experimentales o errores conceptuales en teorías de mecánica coclear. Estos pueden incluso sugerir mecanismos ocultos pasados por alto o no vistos en la cóclea. El modelo permite trabajar con varias estructuras y parámetros en una manera que no podría ser posible en una cóclea real. Los modelos no toman en cuenta los problemas inherentes al trabajo con tejidos biológicos, especialmente en especies vivas. Sin entrar en un tratamiento detallado de cualquier modelo en particular, será útil discutir brevemente algunos de los problemas que los constructores de modelos intentan direccionar. Los primeros modelos físicos y matemáticos de la cóclea, fueron diseñados para reproducir la mecánica coclear y descubrir su esencia. Un buen ejemplo es el modelo de Juergen Tonndorf`s, el cual fue similar al estudiado originalmente por Bekesy. Una membrana de ancho variable separaba dos canales de fluido; este modelo era varias veces el tamaño de la cóclea real. A su vez, los modelos matemáticos realizados, como el de Josef Zwislocki, fueron unidimensionales y se comportaban mejor para frecuencias de entrada con longitudes de onda relativamente largas comparadas a la dimensión transversal de la cóclea. En conjunto, las ondas viajeras producidas en estos modelos relativamente simples imitan aceptablemente el fenómeno observado por Bekesy en cócleas reales de cadáveres humanos. De cualquier manera las décadas pasadas de investigación
han demostrado claramente que los cambios post-mortem en los tejidos cocleares
tienen efectos sustanciales en la mecánica de la sección
coclear. En la cóclea viva se piensa que ocurre un efecto de resonancia
en el peak de la onda viajera, demostrado básicamente por la agudeza
del peak del patrón de desplazamiento. Por lo tanto, ni en las observaciones
de los primeros modelos cocleares, ni las observaciones de Bekesy en cadáveres,
pueden describir adecuadamente el movimiento de la membrana basilar en
una cóclea viva. Efectivamente, el consenso en la actualidad es
que los patrones de vibración de la cóclea viva no pueden
ser completamente explicados por mecanismos completamente pasivos.
|
| Home | Oído Externo | Oído Medio |