| Is the poor growth of maize in the Maputaland area of South Africa caused by mineral deficiencies? |
|
Plan for the experiment:
Results:
Possible reasons:
Impact on the people of the Maputaland area:
Possible future experimentation:
|
5. Possible reasons:
5.1 Reasons for stunted growth and deformities in the Maputaland and deficient solutions:
Nitrate deficiency:
Concentrations of nitrate =
Complete solution: 274 mg/l
Maputaland solution: 8.19 mg/l
This is one of the most significant deficiencies in the Maputaland solution.
The "R" group is the part of the amino acid that differentiates it from other amino acids and gives it its qualities. It is also the part of amino-acids that sulphur is often part of. These amino-acids then form chains which inter-link several to form proteins. Anything that includes protein includes nitrogen therefore it will be found in almost all cell structures. Nitrogen also forms part of many pigments and co-enzymes. It is part of the porphyrin rings which are the basic structure of electron carrying and exciting mechanisms such as chlorophyll and cytochromes. Because nitrogen is so integral to life nitrate deficiency is one of the most common ones, often leading to death and stunting of plants. Surprisingly the nitrate deficient plants grew better than the ones at Maputaland levels. Sulphur deficiency: Sulphate concentration = Complete solution: 78 mg/l Maputaland solution: 28 mg/l This is not as large a deficiency as is seen with the nitrate, yet it may be large enough to cause major stunting. Sulphur is an important part of proteins. It forms part of most amino acids and forms disulphide bridges which hold a protein’s tertiary structure together. Sulphydryl groups form bonding sites in the active sites of many enzymes. (Street & Öpik, 1984) Membrane sulpholipids form part of cells’ recognition signals. Sulphur deficiency’s main symptom is chlorosis in young leaves. Magnesium deficiency: Complete solution: 20 mg/l Maputaland solution: 5 mg/l Magnesium’s main function in plants is as part of the chlorophyll molecule and is also a coenzyme. It activates dehydrogenases and phosphate transfer enzymes. These enzymes are a vital part of an organism’s metabolism so a deficiency can cause an energy deficiency if not totally efficient. (Street & Öpik, 1984). Magnesium, with calcium, is part of pectates that act as glue binding cellulose microfibrils together. (Fogg, 1963) This holds cell walls internally and also attaches adjacent cells to each other. A group of enzymes called kinases need magnesium for activity. These transfer phosphoryl groups from ATP to an acceptor. One of the important kinases that do this is hexokinase which transfers a phosphoryl group from ATP to a variety of hexose sugars at the beginning of glycolysis. (Stryer, 1981) This is why magnesium deficiency often leads to chlorosis and deformed and stunted growth. Iron: Complete solution: ? mg/l (a few drops of FeCl3 were used to provide an excess) Maputaland solution: >0.006 mg/l Though the concentration was not specified in the complete solution if it is compared with the concentrations of other minor essential minerals it seems to be very low in the Maputaland solution. Iron is used in the synthesis of chlorophyll in a precursor to the completed form. Iron is also a part of many of the components of metabolism, namely cytochrome c, cytochrome oxidase and a variety of catalases. (James, 1973) These cytochrome has a similar shape to the chlorophyll molecule shown here but has iron instead of the magnesium. It is also part of iron-sulphur compounds, for example, quinones, these also carry electons in the electon transform chain (Alberts et al, 1989). As such an iron deficiency leads to chlorosis of young leaves and the slowing of metabolic reactions due to the reduced number of iron containing catalases and cytochromes. Potassium deficiency: Complete solution: 239 mg/l Maputaland solution: 12 mg/l Potassium is an essential mineral for plants yet its role is not fully known. It is the most common cation of protoplasm and as such balances the charges on cytoplasmic anions. In doing this potassium is used to maintain turgor and in movement, such as the opening and closing of stomatal guard cells. Chemically sodium has almost identical capabilities and will be used instead of potassium if it is not available yet plants show a preference accumulating potassium. Street and Öpik (1984) say: "The only known metabolic role for potassium is as an activator of some enzymes, but other elements which act as enzyme activators are required in micronutrient quantities only. The affinity of proteins for potassium is, however, low, so that fairly high potassium concentrations may be needed to make potassium-enzyme complexes." Potassium deficiency’s most common symptoms are yellow edges and tips of leaves (Roberts et al, 1993). This was seen in a number of the plants in the Maputaland solution. Potassium is a component of wood ash, which is supplied to the maize in the area by slash and burn agriculture. This suggests that potassium may be an important deficiency of the soil as maize in the area grows best in these burnt fields. Calcium deficiency: Complete solution: 95 mg/l Maputaland solution: 12.5 mg/l Like magnesium calcium forms part of pectates which bind cellulose cell walls together. It also associates as Ca2+ ions with cell membrane phospholipids and is necessary for the maintenance of the permeability of the plasmalemma. Calcium also acts as an activator for some enzymes, namely amylases, ATPases and phospholipases. (Street &Öpik,1984) This should lead to structural deformities and to a breakdown in the transmission of energy Silicon deficiency: Complete solution: 0 mg/l Maputaland solution: 0 mg/l As a grass maize requires silicon for part of its structure, to provide it with rigidity. Silicon deficiency in grasses often leads to wilting, necrosis, and withering of leaves. (Street &Öpik, 1984) These symptoms were seen in the plants and assumed to be symptoms of a disease as it seemed to spread. None of the nutrient solutions or the growth medium contained silicon. Silicon is one of the few abundant minerals in the Maputaland sands so in real life there would not be this deficiency.
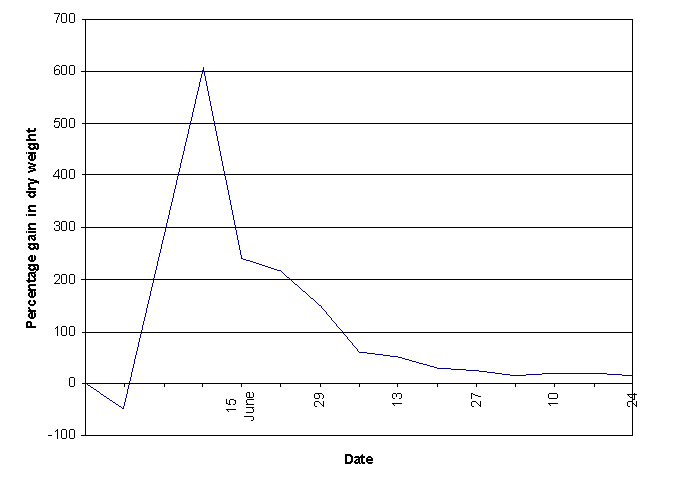
Phosphorus deficiency: Phosphate concentration = Complete solution: 140 mg/l Maputaland solution: 1.6 mg/l in the solution (unknown concentration in soil) Phosphorus is part of nucleic acids and phospholipids (the basic molecule of cell membranes). As part of the adenosine phosphates (ATP, ADP and AMP) the phosphate group takes part in energy transduction. Intermediary metabolism includes phosphorylation, the addition of phosphate groups from ATP to carbohydrates and their intermediary products. In the light dependent photosynthesis ATP is made then used in the Calvin cycle in the production of hexose sugars. As ATP is constantly made and used within cells there is a constant conversion of inorganic phosphate to organic forms and visa versa. (Street & Öpik, 1984) A shortage of phosphorus means a breakdown of the transmission of energy in plants and as such prevents growth. If graph 7 (growth rates in the different nutrient solutions in This factor could either be overcrowding due to the fact that there were five maize plants in each pot or the wilting disease described earlier. The weekly growth rate of maize under natural conditions (simplified after Street & Öpik, 1984)
. . . . . . . . . . . . . . One thing which can be conclusively seen from graph 7 is that there were a number of
external factors which had considerable effects on the growth rate. The growth rate in all solutions apart from the Maputaland one fluctuates on the same days, often by similar amounts. It is well known that small fluctuations in light intensity and temperature can have significant effects on the growth rate of maize (Street & Öpik, 1984). As they are all affected similarly it can be assumed that the fluctuations are not caused by the mineral deficiencies.
5.3 What caused the different germination rates? Magnesium is a cofactor for many enzymes. Some of these enzymes may be vital in the process of germination. For example, hexokinase, which carries out phosphorylation at the beginning of glycolysis, requires magnesium. If this enzyme does not function correctly then respiration will slow down, possibly not providing enough energy for the process of germination. Gibberellins are a group of plant growth substances which are involved in the breaking of dormancy in seeds and in mobilising food reserves during germination. It may be that certain minerals are needed during the synthesis of gibberellins, either as cofactors or as part of enzymes. Phosphorylation occurs a number of times in this synthesis (Devlin & Witham, 1983). This uses kinases which normally require magnesium to do this. ( The different nutrient solutions may have lead to different concentrations, which would affect the rates of osmosis. As a significant part of germination is the absorption of water this can affect the rate. It also will affect the rate of most chemical reactions within the seed/seedling. 5.4 What about the plants native to the Maputaland area? Due to the high water table (usually less than 5 metres from the surface) the Maputaland area has some of the most luxuriant growth for many kilometres. Why are these plants not affected by the mineral deficiencies? What adaptations do they have that cannot be used by maize? Maize, like many grasses is a C4 plant. This means it stores carbon dioxide as sucrose during the night then releasing it again in the day to provide chloroplasts with high CO2 concentrations for photosynthesis. This has the advantage of allowing the plant to have its stomata closed in the heat of the day to prevent water loss while allowing photosynthesis to occur when the light intensity is greatest. It has to provide this stored carbon dioxide to prevent concentrations of the gas falling far enough to cause a reaction called photorespiration occurring which reverses the flow of CO2 in the fixation process. (Alberts et al, 1989) This process may mean that maize has different mineral requirements to non-C4 plants. This is unlikely to be the reason for their ability to grow in the area because other grasses, presumably also C4 plants, grow reasonably well in the sands. Another reason may be that most of the area’s plants are specifically adapted to the local conditions. This is a possibility as a large number of species are endemic to the area. This is especially true of the sandforest and dune-forest tree species. 5.5 Conclusions: Sulphur, magnesium and nitrate are equally important to maize during the phase of growth measured in the experiment, though the plants’ requirements may change as they grow. Due to leaching of minerals from the sand there is a general deficiency of all major essential minerals and probably also of minor essential minerals. There is a prominent deficiency of nitrate which probably is the greatest cause of stunted growth.
|
|
|