Сибаров Д.А., Коваленко Р.И., Ноздрачев А.Д.,
Особенности функционирования пинеалоцитов крыс при стрессе в светлое время суток
Росс.Физиол.журн., 2000, 86(8):1049-1052.
Кафедра общей физиологии, Санкт-Петербургского государственного университета, Россия, 199034, Санкт-Петербург, Университетская наб. 7/9.
В дневное время проводилось электрофизиологическое исследование эпифизов крыс - интактных и подвергнутых 48-часовой водной и пищевой депривации (осмотический стресс). Анализировалась частота внеклеточно отведенных разрядов пинеалоцитов. Обнаружено наличие в эпифизе нескольких типов клеточной активности, что согласуется с результатами других исследователей. Частота разрядов пинеалоцитов у животных при стрессе была в 4-6 раз выше, чем у интактных, за сче т увеличения доли "быстрых" клеток (>5 имп/с), а также перехода части пинеалоцитов с регулярного на пачечный тип активности. Электрическая стимуляция обонятельных луковиц снижала частоту разрядов наиболее активных пинеалоцитов. Усиление электрической активности клеток эпифиза отражает увеличение интенсивности экзоцитоза клетками эпифиза веществ белково-пептидной природы и, вероятно, серотонина, но не мелатонина Блокирование экзоцитоза колхицином выявило тесную связь секреторной активности пинеалоцито в в дневное время с их электрическими разрядами. Показано существование центральных механизмов ограничения активности пинеалоцитов со стороны, в частности, обонятельных структур.
Ключевые слова: эпифиз, пинеалоциты, электрофизиологическое исследование, осмотический стресс, секреция
Эпифиз, как фотонейроэндокринный орган, модулирует активность многих физиологических систем в связи с циркадными ритмами. Он участвует в управлении адаптивными реакциями организма на изменение внешних у словий среды и внутреннего гомеостаза [2,10,14]. Долгое время считалось, что единственной функцией эпифиза, расположенного вокруг бухты III-го мозгового желудочка, является сезонная и суточная модуляция деятельности репродуктивной системы. Однако более п оздние исследования показали участие эпифиза в стрессорных и иммунных реакциях. Наряду с гипоталамо-гипофизарным комплексом, пинеальная железа вовлекается в формирование множественных адаптационных ответов организма на стрессорное воздействие, главным об разом, за счет влияний со стороны краниального шейного ганглия и ЦНС в I-ю фазу общего адаптационного синдрома и за счет действия гормонов коры надпочечников во II-ю его фазу [1]. Показано, что под влиянием страха, шума, вибрации, иммобилизации, гипергли кемии, отсадки потомства у лактирующих животных, принудительного купания в холодной воде усиливается продукция эпифизом мелатонина и веществ пептидной природы [1,2]. В частности, мелатонин (основной гормон эпифиза) ослабляет негативное воздействие на орг анизм окислительного, осмотического, психического и других видов стресса [1,2]. Этот нейрогормон также обладает антиканцерогенными свойствами и может модулировать функции иммунной системы [7,18]. Выделенные из ткани эпифиза пептиды проявляют сходные с ме латонином эффекты. Пептидные препараты эпифиза способны повышать функциональную активность тимуса, нормализовать ряд нарушений углеводно-жирового обмена, обладают антиокислительными свойствами [3,9]. Нами сообщалось [5], что при осмотическом стрессе прои сходит активация синтеза и экскреции веществ белково-пептидной природы одним из типов клеток эпифиза - светлыми пинеалоцитами.
Известно, что микроэлектродная регистрация потенциалов действия позволяет оценить уровень секреторной активности пинеалоцитов, поскольку деполяризация нейросекреторных клеток коррелирует с экзоцитозом [4,16]. В связи с тем, что жирорас творимый мелатонин способен диффундировать через клеточную мембрану, его секреция, по-видимому осуществляется не путем выведения секреторных везикул и, следовательно, напрямую не связана с деполяризацией мембраны. Это, в частности, подтверждается следую щими наблюдениями. Подавление потенциалов действия пинеалоцитов с помощью Ca2+-связывающих веществ верапамила или нифедипина не оказывает существенного влияния на продукцию мела тонина [13], а угнетение активности N-ацетилтрансферазы (фермента, катализирующего синтез мелатонина) не влияет на электрическую активность пинеалоцитов. Полагают, что секреция же эпифизарных пептидов осуществляется обычным экзоцитозом.
Целью данной работы была проверка ранее полученных нами данных об активации пинеалоцитов при осмотическом стрессе при помощи методики регистрации частоты их деполяризации. Кроме того, одной из задач явилось изучение влияния на эпифиз с о стороны обонятельной системы, поскольку у крыс она имеет даже большее значение, чем зрительная.
МАТЕРИАЛЫ И МЕТОДЫ
Объектом исследования служили 16 белых крыс-самцов линии Wistar с массой тела 160-180г, содержавшихся в виварии СПбГУ при естественном освещении. Опыты проводились в светлое время суток 12:00 - 13:00. К онтролем служили 4 интактных животных, имевших свободный доступ к воде и пище. Крысы опытной группы (N=12) подвергались 48-часовой водной и пищевой депривации. Электрическая активность клеток оценивалась при помощи микроэлектродной регистрации разрядов п инеалоцитов. Наркотизированные уретаном крысы (1.1 г/кг массы тела, внутрибрюшинно) помещались в стереотаксический прибор. Производилось удаление фрагмента кости черепа (диаметром 5мм) в области брегмы, перевязывался венозный синус. Внеклеточная регистр ация биопотенциалов осуществлялась при помощи заполненного 3М NaCl стеклянного микроэлектрода (диаметр кончика 10-30 мкм, сопротивление 4-6 МОм). Сигнал с микроэлектрода усиливался и затем оцифровывался при помощи звуковой платы “Sound-Blaster 16” под у правлением программы “Cool-Edit Pro”. После записи спонтанной клеточной активности эпифиза каждая крыса подвергалась электрической стимуляции обонятельного эпителия при помощи двух серебряных биполярных электродов, которые вводились в носовые ходы на глу бину 5-6 мм (параметры раздражающего тока: 40В, 100Гц, 50 мсек в течение 15сек). Микроэлектродная регистрация разрядов пинеалоцитов производилась непрерывно до, во время и после стимуляции. В двух отдельных экспериментах для выяснения связи между электр ической активностью клеток и экзоцитозом секреторных гранул, в микроэлектродный раствор добавлялся колхицин в концентрации 0.01М, блокирующий разборку микротрубочек и процессы экзоцитоза в клетке.
Статистическая обработка включала программный поиск отдельных разрядов в мультиклеточной записи и идентификацию разрядов отдельных клеток в разработанной нами программе “SMP 5.0” следующими способами: построение гистограммы распределен ия амплитуд отдельных разрядов; разделение разрядов по полярности; построение совместного распределения по амплитуде, продолжительности де- и реполяризации; вычисление автокорреляционной функции для поиска периодических процессов и зависимостей между раз рядами клеток; наложение множества периодических промежутков равной длины для выявления периодических процессов; поиск межклеточных взаимодействий путем наложения множества промежутков, начинающихся разрядом с заданными параметрами, для выявления отвечаю щих на него разрядов других клеток.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
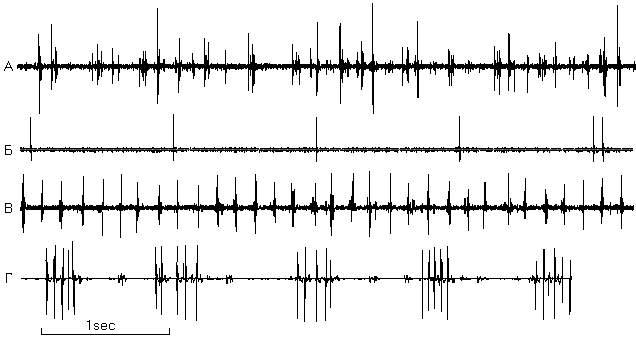
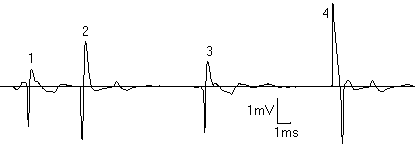
В ходе экспериментов обнаружены "медленный", с частотой разрядов < 2 имп/с, и "быстрый", с частотой разрядов >4 имп/с, типы активности пинеалоцитов. "Медленный" тип был представлен регулярными, не регулярными и пачечными разрядами (Рис.1). Электрофизиологичекие параметры активности клеток у интактных и подвергнутых осмотическому стрессу животных представлены в таблице 1. В эпифизе интактных крыс наблюдался, как правило, только "медленный" тип акт ивности (частота 0.86±0.49 имп/с, регулярно и нерегулярно разряжающиеся клетки), что свидетельствует о невысокой секреторной активность эпифиза у интактных животных.
Ранее мы сделали тот же вывод на основании электронно-микроскопических исследований, в которых оценивался объем везикул различной природы в эпифизе интактных крыс [8]. В опытах с добавлением в микроэлектродный раствор колхицина, блокир ующего разборку микротрубочек и, в результате этого, останавливающего процессы экзоцитоза в пинеалоцитах, мы получили снижение электрической активности клеток, прилежащих к кончику микроэлектрода. Это наблюдение свидетельствует о существовании связи разр ядов пинеалоцитов с выбросом ими секреторных гранул.
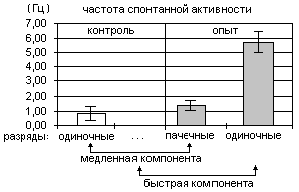
Регистрация разрядов пинеалоцитов у крыс, подвергавшихся осмотическому стрессу, выявила клетки с пачечным типом активности, отсутствовавшие у интактных животных (Рис.2). Переход при стрессе части клеток с "медленным" типом активности с регулярного на пачечный тип разрядов приводил к увеличению суммарной частоты разрядов в 4-6 раз. Кроме того, появлялся "быстрый" тип активности (5.71±0.73 имп/с).
Сделанное нами предположение о переключении части пинеалоцитов с регулярного на пачечный тип активности подтверждается следующими наблюдениями: обратимостью перехода с регулярного на пачечный тип разрядов отдельных клеток; переменным ч ислом разрядов в пачке; близостью частот разрядов и частот пачек у клеток с "медленным" типом активности, а также близостью продолжительности отдельных разрядов у пачечно и регулярно разряжающихся клеток. Наши данные о существовании нескольких типов разр ядов пинеалоцитов согласуются с наблюдениями Reyes-Vazquez [20]. Эти наблюдения о существовании нескольких типов клеток представляются весьма интересными, так как долгое время в эпифизе различали только 2 типа пинеалоцитов - "светлые" и "темные". Недавно с помощью световой микроскопии идентифицировано большее число типов клеток: светлые, темные, промежуточные и гранулярные [6]. По мнению автора, принадлежность клетки к определенному типу определяется ее текущим функциональным состоянием. Пинеалоциты с р егулярным, нерегулярным и пачечным типами разрядов изучались многими авторами [17,19,20], и было установлено, что продолжительность отдельных разрядов может варьировать от 1 до 200 мс. Пос леднее значительно облегчает идентификацию разрядов отдельных клеток по форме при мультиклеточном отведении.
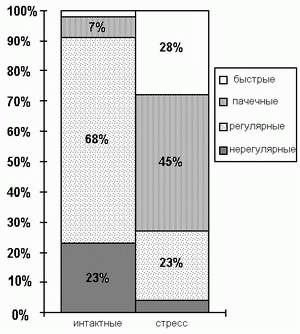
Соотношение пинеалоцитов с вышеуказанными типами активности различалось у интактных и подвергнутых осмотическому стрессу крыс (Рис.3). Основная часть клеток интактных животных (91%) была представлена редко разряжающимися "регулярными" и "нерегулярными" клетками, а при осмотическом стрессе возрастала доля пинеалоцитов с "пачечным" и "быстрым" типами активности, что приводило к росту суммарной частоты разрядов. Одновременное 3-кратное снижение числа "регулярных" и 6,5-кратный рост числ а "пачечных" клеток также указывают на переключение клеток на пачечные тип разрядов. Последнее вместе со снижением числа медленно разряжающихся "нерегулярных" клеток и появлением пинеалоцитов с "быстрым" типом активности указывают на интенсификацию экзоц итоза в эпифизе у животных, подвергавшихся осмотическому стрессе. Наблюдаемую активацию секреторных процессов в клетках эпифиза можно обьяснить тем, что увеличение осмолярности крови в капиллярах эпифиза, вызванное 48-часовой водной и пищевой депривацией , сопровождается усилением выхода воды из его клеток. Выход воды из пинеалоцитов вместе с секреторными продуктами, возможно, происходит в результате увеличения в крови концентрации глюкокортикоидных гормонов, характерного для II-стадии общего адаптационн ого синдрома. Как известно, последние подавляют секрецию инсулина, что приводит к увеличению содержания глюкозы в крови и повышению ее осмолярности [10]. Кроме того, пинеалоциты имеют рецепторы к глюкокортикоидам, активация которых приводит к усилению се креторных процессов в клетках [2].
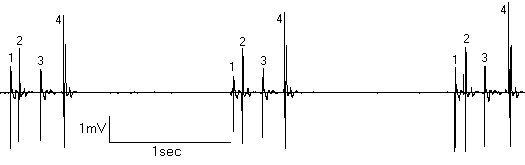
В условиях активации электрических процесов в эпифизе у депривированных животных нам удалось наблюдать группы взаимодействующих пинеалоцитов (рис.4). Внешне активность такой группы напоминает пачку, но клетки различаются по форме спай ков и разряжаются в строгой последовательности с различными, но постоянными задержками между спайками. Подобные межклеточные взаимодействия, возможно, обеспечивают активацию и синхронную деятельность целой группы пинеалоцитов за счет существования плотных [15] и щелевых [8] межклеточных контактов.
Прямые эфферентные нервные связи обонятельных структур с эпифизом были продемонстрированы многими методами. Например, после разрушения хабенулярного ядра в эпифизе выявлялись дегенерирующие нервные волокна [21]. Еще более убедительно т акие связи были показаны в электрофизиологических [11,21] и гистохимических работах [12,22]. Даже полная симпатоэктомия эпифиза не блокирует вызванные потенциалы в пинеалоцитах при электростимуляции габенул. Нами обнаружено, что электрическая стимуляция обонятельного эпителия приводит к достоверному снижению суммарной частоты разрядов клеток эпифиза на 58% у депривированных и на 45% у интактных крыс. Главным образом, при стимуляции обонятельного эпителия снижалась частота разрядов "медленных" пинеалоци тов. Клетки с "быстрым" типом активности на нее практически не реагировали. Это еще раз подтверждает положение о гетерогенности клеточных популяций пинеалоцитов в эпифизе и говорит от том, что обонятельный сенсорный вход контролирует деятельность только части клеток эпифиза.
В целом, нами впервые получены данные об активации части клеток эпифиза в дневное время у крыс при осмотическом стрессе, обнаружена ранее не описанная в литературе взаимная активация пинеалоцитов путем межклеточных взаимодействий. Пока зано также ограничивающее влияние на частоту разрядов наиболее активных пинеалоцитов со стороны обонятельных структур.
СПИСОК ЛИТЕРАТУРЫ
1.
Анисимов В.Н. Физиологические функции эпифиза (геронтологический аспект)
Росс. Физиол. Журн. 83(8): 1-13. 1997. РИСУНКИ
2. Аpушанян Э.Б., Аpушанян Л.Г., Элькебян К.С. Место эпифизаpно - адpенокоpтикотpопных
отношений в попpавочной pегуляции поведения Успехи физиол. наук.24(4):
12-23. 1993.
3. Галанцев В.П., Коваленко Р.И., Камардина Т.А., Попов С.М., Январева
И.Н., Перепелица В.А. Влияние пептидов эпифиза на резистентность сердца
и состояние тканевых систем перекисного окисления липидов у крыс при асфиксии
и реоксигенации Вестник СПбГУ, сер.3, 3(17): 68-78. 1995.
4. Глебов Р.Н. Эндоцитоз и экзоцитоз М., Высшая школа, 95с. 1987.
5. Коваленко Р.И., Сибаров Д.А., Павленко И.Н., Лукьянова Е.Л. Структура
пинеалоцитов крысы при стрессе и после унилатеральных интраназальных введений
окситоцина Росс. Физиол. Журн., 83(8): 78-93.1997.
6. Колесникова Л.А. Эпифиз относительно диких и доместицированных лисиц:
морфофункциональные изменения в течение суток Физиол. Журнал. им. И.М.Сеченова
82(2): 91-97. 1995.
7. Морозов В.Г., Хавинсон В.Х. Пептидные биорегуляторы (25-летний опыт
экспериментального и клинического изучения) СПб. Наука, 74с. 1996
8. Скопичев В.Г., Коваленко Р.И., Селиверстов Ю.А. Изменение ультраструктуры
пинеалоцитов крысы в отдаленные сроки после гипофизэктомии Цитология,
29(5): 516-520. 1987.
9. Слепушкин В.Д., Пашинский В.Г. Эпифиз и адаптация организма Томск,
изд-во Томского ун-та, 210с. 1982.
10. Чеpнышева М.П. Гоpмоны животных.- СПб.,Глаголъ 296с. 1995.
11. Demaine C., Semm P. Electrophysiological evidence for central nervous
connections of the pigeon's pineal gland Brain Res. Bull. 13(5):629-634.
1984.
12. Jimenez A.J., Fernandez-Llebrez P., Perez-Figares J.M. Central projections
from the goldfish pineal organ traced by HRP-immunocytochemistry Histol.
Histopathol. 10(4):847-852. 1995.
13. McCance I., Parkington H.C., Coleman H.A. The association between
melatonin production and electrophysiology of the guinea pig pineal gland,
J.Pineal Res. Sep;21(2):79-90.1996.
14. Milin J.R. Morphodynamic response of the pineal gland to initial stress
attack Arch. d'Anat. micr. 73(3): 159-180. 1984.
15. Kunwar P. The ultractructure of mammalian pinealocytes: a systematic
investigations. Micr.Res.and Tech., 21 : 85-115.1992.
16. Nordmann J.J. (1983) Stimulus - secretio coupling Progr. Brain. Res.
60: 283-303
17. Parkington H.C., McCance I., Coleman H.A. Two types of cells with
central innervation in pineal gland of guinea pigs, Am.J.Physiol, 252
: 369-377. 1987.
18. Reiter R.J., Melchiorri D., Sewerinek E., Poeggler B., Barlow-Walden
L., Chuang J., Ortiz G.G., Acuana-Castroviejo D.A. Review of the evidence
supporting melatonin’s role as an antioxidant J.Pineal Res., 68: 1-11.
1995.</ LI>
19. Reuss S., Semm P., Vollrath L. Changes in the electrical activity
of the rat pineal gland following stimulation of the cervical sympathetic
ganglia, J.Autonom Nerv. Syst. Apr;12(4):281-288.1985.
20. Reyes-Vazquez C., Prieto-Gomez B., Aldes L.D., Dafny N. Rat pineal
exhibits two electrophysiological patterns of response to microiontophoretic
norepinephrine application, J. Pineal Res. 3(3) : 213-222.1986.
21. Ronnekleiv O.K., Kelly M.J., Wurrke. W. Single unit recordings in
the rat pineal gland: evidence for habenular-pineal neural connections,
Exp. Brain Res., 39 : 187-192.1980.
22. Yanez J., Anadon R. Afferent and efferent connections of the habenula
in the rainbow trout (Oncorhynchus mykiss): an indocarbocyanine dye (DiI)
study J. Comp. Neurol. 372(4):529-543. 1996.
Daytime rat pinealocytes function in stress
D.A.Sibarov, R.I.Kovalenko, A.D.Nozdrachev.
General physiology department, St.Petersburg State University,
Russia, 199034, St.Petersburg, University emb. 7/9.
Daytime electrophysiological study of pineal gland was made in intact and in osmotic stress subjected rats (48-hours water and food deprivation). Frequency of extracellularly recorded spikes was analyzed. Several types of cellular elec trical activity were detected in the pineal gland, which is in agree with other investigators results. Total frequency of pinealocytes discharging in stress was 4-6 times higher, than in intacts, due to an increase of "fast" cells (>5 imp/sec) number and due to switching of some cells from regular to pattern type of activity. Electrical stimulation of olfactory epithelium decreased the frequency of spikes of the most active pinealocytes. Daytime increase of pineal electrical activity reflects the in tensification of secretion of protein/peptide substances and, probably, of serotonin, but not of melatonin. Blocking of exocytosis with colchicin revealed the tight link of pinealocytes secretion with their electrical activity. An existence of central (olfactory, in particular) mechanisms, limiting the pineal activation was shown.
Keywords: pineal gland, pinealocytes, electrophysiological study, osmotic stress, secretion