The Environmental Effects of Dams
Rivers are highly dynamic systems where any one impact may have far-reaching ramifications. The interdependence of biological and physicochemical processes makes distinctions between these types of effects difficult, and consequently these categories are not used in this report. Instead, this paper attempts to identify the direct effects of damming rivers, and then explores the cascade of indirect effects that result. This method acknowledges the complexity of the real world even though it can lead to some overlap and redundancy where direct effects lead to similar indirect impacts.
Dams as Physical Barriers
Dams block the downstream passage of water so that the upstream water level rises, and a lake or impoundment forms. In the process, dams also prevent the free movement of aquatic plants and animals past the obstacle formed in the river.
Upstream Migration of Adult Anadromous Fish
It is well-known that dams block the upstream migration of anadromous fishes such as Salmon and Shad (e.g., Yoshiyama et al., 1998; Moser et al., 2000; Smith et al., 2000), and even some invertebrates (Fičvet, 2000). This has particular importance for anadromous species because they must bypass dams to reach river headwaters to reproduce in suitable spawning habitat.
Runs of returning adult fish are often important resources for wildlife. For example, grizzly bears depend on protein-rich roe from salmon to develop fat reserves to survive hibernation. Recently, it has been shown that nutrients from the carcasses of spawning salmon also help fertilize vegetation near rivers, and this has cascading effects through the terrestrial food chain (Ben-David et al., 1998). Without this nutrient resource, terrestrial production will suffer, particularly if nitrogen is a limiting nutrient for plant growth in the system.
If adult fish cannot progress upstream then affected populations fish are unlikely to persist because most anadromous fish species are drawn only to spawn in their natal rivers. When confronted by impassable obstacles, adult salmon, for example, do not return to sea to find other rivers. Instead, they remain trapped below the obstacle making repeated efforts to travel upstream. This can result in highly emotive scenes where dead and dying salmon collect in backwaters and pools. Native Americans in the Port Angeles area of Washington State still recall images of hundreds of thousands of salmon dying in pools below the Glines Canyon Dam in the years following its construction (DOI, 1996). Even where dams are very low head they can provide significant obstacles to upstream migration. The Quaker Neck Dam in North Carolina, for example, has been shown to be a significant barrier to striped bass and American shad even though it can be completely submerged by higher flows (Beasley and Hightower, 1998).
Some species of affected fish have attempted to adapt to the presence of obstacles by spawning below offending dams (Zhong & Power, 1996; Dauble et al., 1999). However, there is no evidence that this strategy has been successful. Indeed, Dauble et al. showed that the number of redds below dams on the Snake River (WA) declined from 12% of the total annual spawning activity in 1994 to less than 5% in 1997. The successful production of larvae quickly declines where suboptimal physical conditions exist (Maret et al., 1992). Suitable spawning habitat usually exists in the headwaters and tributaries of rivers, not the areas below dams, making it unsurprising that this reproductive strategy has resulted in poor viability of redds.
Human response to the plight of returning anadromous fishes has, in some cases, been the installation of structures that allow fish to pass over the dams. Fish ladders and other similar structures have undergone extensive research and development to maximize the number of adults able to successfully navigate them. In some situations, this technology has even been applied to natural obstructions in rivers, such as waterfalls, to increase the spawning habitat available to adult fishes (Bryant et al., 1999). However, the usefulness of fish ladders is strongly dependent on the species trying to bypass a dam. While many salmonid species are able to use fish ladders, some species such as White Sturgeon, Alewives, Striped Bass, and American Shad, either cannot use them or do so in only small numbers (Anon, 1999; Bowman & Hightower, 2000). Even where fish ladders are successful in helping fish to bypass smaller dams, the effort required to negotiate passage over tall dams can be exhausting. Exhausted fish may be more susceptible to natural dangers such as predation and disease, and are also more likely to be caught up in strong surface currents such as those that result from water flowing through dam spillways. PATH (1999) estimates that up to 40% of adult fish that successfully climb fish ladders over high-head dams are caught up in spillway currents and swept back over the dam to the river below.
A less common alternative to installing fish ladders is to physically capture the adult fishes and transport them above dams using purpose-built tankers. However, this procedure has inherent problems such as stress and damage from capture and handling that result in fish deaths during transport, and indirect or delayed mortalities that happen after release.
Non-seafaring Species
Problems associated with the upstream migration of anadromous (and diadromous) species receive a great deal of attention, but the fragmentation of river systems also has implications for species that reside within rivers for their entire life history. For example, Smallmouth Bass (Micropterus dolamieu) spend most of their life in a small stretch of a river but make significant migrations along the river in order to spawn (Kanehl et al., 1997). Clearly, dams prevent the migration of these species as well as the more publicized anadromous species. Similarly, Hildebrand & Kershner (2000) estimate that the minimum uninterrupted length of river required to support Cutthroat Tout populations is 9.3km for high-density populations and 27.8km for low-density populations. The minimum stream length raises issues of sustainability in some areas where rivers are dammed repeatedly. For example, Kanehl et al. calculate that there is one dam for every 14.5 km of flowing water in Wisconsin. It may be that ignorance of the requirement for uninterrupted river length was instrumental in the extirpation of four minnow species species in Oklahoma, which resulted from damming small prairie streams (Winston et al., 1991).
The implications are somewhat less severe for insects with aquatic larval stages because adults can fly past the dam to recolonize reaches of river upstream of dams. However, other aquatic invertebrates such as crustaceans and molluscs are unable to recolonize areas above dams if upstream populations are lost.
Exotic Fish Management
The fact that dams act as migration barriers to fish does have some positive implications for the management of exotic species that invade river systems. For example, the invasion of parasitic Lampreys into the Great Lakes of North America has badly affected native populations of salmonids that are not adapted to the presence of Lampreys. Low-head dams have been shown to be effective in preventing the further spread of Lampreys into some river tributaries that form part of the system (Porto et al., 1999). Similarly, Brook Trout are an introduced species in several midwest and northwest states of the USA. Brook trout have adversely impacted native species through competitive exclusion (e.g. Van Kirk & Griffin, 1997). Artificial barriers (dams) have been used to limit the range of the introduced species in order to maintain populations of native species (Thompson & Rahel, 1998).
Downstream Migration of Larvae and Juvenile Fish
Mortality rates for larval and juvenile fishes are particularly important for determining whether populations are able to replace themselves. Even a slight increase in instantaneous mortality rates for larval fish (and other larval-stage organisms) can have dramatic effects on the number of recruits that survive to the next stage of development (Underwood & Fairweather, 1989). Consequently, extra mortalities associated with downstream passage past dams take on an added significance.
Depending on the species, larval stage fishes either remain in instream habitats while they grow larger or else are swept downstream to arrive in the sea soon after hatching. For example, Chinook salmon spend between 1 and 18 months in the stream prior to migrating to the sea, while pink salmon larvae drift down to the sea immediately after hatching and spend most of their juvenile life in estuarine areas. In the normal course of development, pink salmon larvae are swept down the river by strong water currents and arrive at the sea within a few days of hatching. However, when a dam is built velocities greatly decrease because a lake forms above the dam. Without strong currents to sweep them downstream, the journey to the sea becomes greatly extended for outmigrating larvae and juveniles. This is particularly important for larvae of pink salmon because they cannot swim effectively. The larvae must reach the estuary reasonably soon or else they will starve in the absence of suitable planktonic prey. Also, there is a massive physiological adjustment required to change from a freshwater existence to a marine one, consequently the timing of the arrival at sea is doubly critical for pink salmon larvae.
Wind often generates the currents in lakes and these ‘false’ currents may mislead juvenile fishes that are seeking the lake outlet, further delaying outmigration. Even close to the dam itself, outmigrating juveniles can get disoriented in the forebay and can experience significant delays (Veditti et al., 2000). Delays during this part of the life history can be indirectly fatal because of increased predation rates in the clear waters of the lake (Gregory & Levings, 1998) as well as increased exposure to parasites and disease that flourish in the warm, still, surface waters of lakes (DOI, 1994).
Once larvae and juveniles arrive at the dam itself, they must either pass through the turbines or else over spillways in order to reach the sea. Both present their own set of hazards for the developing fishes.
Spillways can cause significant mortality rates for fish that attempt this route. The fall to the river below, particularly for very tall dams, can stun or kill fish. Bickford and Skalski (2000) report spillway-related mortality rates of up to 18% for juvenile Chinook Salmon. Spillways also create the phenomenon of gas supersaturation. The massive turbulence of falling water entrains air and causes rapid fluctuations in pressure, causing the water to become supersaturated with nitrogen and oxygen. This can cause fishes to develop gas bubble disease, a condition analogous to ‘the bends’ suffered by divers who surface too rapidly. Mortality associated with gas bubble disease depends on both the level and duration of exposure to gas supersaturation. For example, Counihan et al. (1998) found that White Sturgeon larvae suffered a 50% mortality rate after 13 days exposure, but none after 10 days even though signs of gas bubble disease were present after only 15 minutes of exposure to supersaturated water. Ryan et al. (2000) reported that the high variation in experimental results make it difficult to detect relationships between mortality and gas supersaturation. It is possible that gas bubble disease may cause significant sublethal effects, such as reduced growth rates, or increased susceptibility to predation, but this has not been specifically documented yet.
Turbine damage to outmigrating fish has historically been a major concern acknowledged by dam operators (Coutant & Whitney, 2000; Sorenson et al., 1998; Bickford & Skalski, 2000). For example, turbine mortality rates of 10-15% (Anderson et al., 1996) and c. 12% (Bickford & Skalski, 2000) have been reported for salmon juveniles. Attempts to mitigate this include redesigned turbines (with tighter tolerances to reduce the incidence of fish strike), turbine inlet fish screens, and specially designed fish bypass channels that divert fish away from turbine inlets and transport them down to the foot of the dam, or to holding tanks for downstream transportation. The effectiveness of bypass systems for diverting fish away from turbines in hydropower dams varies from dam to dam. Anderson et al. (1996) state that diversion efficiencies of 70-80% would be considered acceptable, but are seldom achieved even in the relatively modern dams on the Columbia and Snake Rivers in the Pacific Northwest of the USA. For example, Johnson et al. (2000) report that the best diversion efficiency for surface bypass devices on the Lower Granite Dam on the Snake River (WA) is 40% of the total number of salmon smolts trying to swim downstream. One way to increase diversion away from the turbines is to increase the flow of water over spillways. However, increased flows over spillways increase the potential for gas supersaturation problems (Anderson et al., 1996).
Once juvenile fish get past the dams they are often weakened and disoriented due to the rigors of passage. Unsurprisingly, predatory fish species have been shown to concentrate their hunting efforts on the tailraces of hydropower dams (Blackwell & Juanes, 1998; Shively et al, 1998; Jepsen et al, 2000).
Overall, juvenile mortalities directly associated with the downstream passage past dams can be very significant. For instance, PATH (1999) estimates that juvenile mortality rates of c. 20% per dam traversed exist for dams in the lower Snake River. Since there are four large dams along the lower part of the river this extra mortality will undoubtedly have massive impacts on the salmon populations in the river. Although dams in other rivers will have different bypass survival rates that are determined by their unique patterns of flow, and also design differences, it is unlikely that figures for the Snake River dams are exceptionally poor because they are relatively modern structures and have been designed to minimize fish impacts.
The Formation of Lakes in River Reaches
Subsidence and Landslides
There may be an increased risk of landslides on slopes that are impacted by increased water levels in reservoirs. Saturation of previously dry soils may cause leaching of soil ions and decrease soil stability.
Changes in Community Structure
Lakes are present on many river systems and provide habitats that some plants and animals require for survival. Lakes also provide opportunities for recreational pastimes that are not available on rivers and are valued for their ability to store water. However, lakes have properties that are sometimes ill-suited for species that are adapted to free flowing rivers and, consequently, the creation of new lakes directly reduces the amount of riverine habitat available. Likewise, artificial lakes may adversely impact species, such as the larvae of pink salmon, that have difficulty traversing the relatively still, open-water conditions found in lakes.
One of the primary results of lake formation is an increase in the abundance of wildfowl and other birds. For instance, the formation of Lake Powell in Colorado has reportedly led to an increase in avian abundance (particularly waterfowl), including the establishment of a significant breeding population of the endangered Peregrine Falcon that was not previously present (Stevens et al., 1997). However, the loss of riverine habitat may affect bird species that are adapted to free-flowing (lotic) water conditions.
When a lake is formed, instream community structure inevitably shifts. This normally means an increase in lentic species such as bloodworms (Chironomidae) and a reduction in lotic species such as stoneflies (Plecoptera) and mayflies (Ephemeroptera) (e.g. Kanehl & Lyons, 1997). For example, 5 species of river-loving fish were eliminated in the Colorado River after creating Lake Powell (Miller, 2000) while introduced species of lake-dwelling fish flourished. Even for generalist species, such as trout, there may be sublethal effects such as differences in growth rates because of inferior food quality in lentic environments. For example, Soluk & Richardson (1997) reported that growth rates of cutthroat trout were reduced when stonefly larvae were not available as a prey item.
Sedimentation of Lakes
Rivers flow relatively quickly and often contain significant loads of suspended sediments. When water velocities are high, the energy and turbulence in the water keeps the sediment particles suspended. However, when water velocities slow down the sediment particles are able to settle with greater ease. Consequently, when rivers enter lakes the suspended load in the river water is dumped on the lake bottom and is seldom re-suspended. Thick layers of sediment provide suitable habitat only for some burrowing invertebrate forms, particularly in deep parts of lakes where light penetration doesn’t allow the growth of aquatic plants or algae. Additionally, organic sediment and debris is slowly decomposed by bacteria which utilize the available oxygen in the water, creating anaerobic conditions near the bottom that are inimical to life (Chatterjee, 1997).
Over time, sediment smothers lake beds and begins to slowly fill the lake in. In cases where the suspended loads of rivers are very high, such as in the Colorado River, lakes may fill quite rapidly. For instance, Lake Powell is the largest man-made lake in the world but is expected to fill with sediment in approximately 500 years (US Bureau of Reclamation, cited on Friends of Lake Powell, 2000). This obviously has long-term implications for the useful life-span of hydroelectric and water storage reservoirs and raises questions about how to ameliorate the rate of lake infilling, and what to do with sediment-filled lakes.
Loss of Sediment and Gravels to Downstream Reaches
Since the sediment in rivers gets deposited in reservoirs, it can no longer reach downstream areas such as estuaries. Ibanez et al. (1998) reported that sediment transport to the Ebro Estuary was reduced by 99% after the construction of a hydropower dam in the lower Ebro River. Consequently, estuarine wetlands start to erode because there are no significant inputs of sediment to balance the erosive processes caused by wave action. The loss of water and sediment due to interception in Lake Powell has been blamed for the failing health of the Sea of Cortez and the delta of the Colorado River (Miller, 2000). Likewise, the Department of the Interior (USA) has recognized that erosion of estuarine and near shore habitat at Port Angeles Harbor (WA) is due to sediment trapping in lakes Mills and Aldwell on the Elwha River (DOI, 1994).
Dams and their reservoirs intercept the downstream migration of riverbed gravels, but this downstream movement continues unabated below the dams. Without replenishment from upstream sources, this leads to an increase in the mean size of riverbed materials (DOI, 1994; Sear, 1995). This has important implications for fish that normally spawn in the main stem of rivers, or those that are forced to spawn below dams, because substrates can become too coarse for successful spawning. For example, Sear (1995) reported an increase in the size of surface gravels below a dam on the River Tyne in the UK. Sear also found that fine sediment berms developed at the extremes of the normal range of flow levels, and that fine material increased in downstream spawning gravels suggesting that downstream siltation is possibly the result of changes in normal flow regimes. The coarsening of riverbed substrates, as well as increased sediment deposition rates, may explain reductions in spawning activity below dams over time, as reported by Dauble et al. (1999).
Accumulation of Toxins in Lake Sediments
The problem of sediment deposition in lakes worsens when toxins such as heavy metals, polychorinated biphenols (PCB’s), or insecticides accumulate in the sediments. This has important implications for future uses that might be made of the sediment, since it is poisonous and potentially unsafe for use as topsoil or landfill. Contaminants in reservoir sediments can persist for decades or centuries, and industrial discharges from industries may still be present in lake sediments long after the industry responsible for the toxins has gone. Sediment contamination also has ecological consequences for aquatic invertebrates, terrestrial insectivores that eat them, and carnivores that devour these in turn. An example of this was reported by Rheinhold et al. (1999) who showed that contaminants from sediments accumulated in larval stage chironomids (bloodworms), and that these toxins were, in turn, accumulated in Pond Bats (Myotis dasycneme) that ate adult chironomids. High levels of toxic compounds in the tissues of animals will have implications for their overall health and reproductive success. The bio-accumulation of toxins through food chains usually impacts most strongly on top-level carnivores, and this principle has been widely reported for species such as swordfish, whales, dolphins, and raptors. A well known example of this was the insecticide DDT that caused the egg shells of raptors to become too fragile to survive. Raptors were affected because DDT was present in the tissues of their prey species.
Turbidity
As another consequence of sediment deposition in lakes, the lake water becomes less turbid than the rivers feeding into them. Because the water tends to be so clear, aquatic plants are able to extend into deep waters because of better light penetration. Without strong currents to reduce plant densities, excessive plant growth may create navigational difficulties (e.g. Toth et al., 1997). Moreover, species that are adapted to river currents may be displaced or eliminated by plants better suited to the still waters of lakes. Consequently, exotic plants can become established and dominant in lakes.
Clear water in lakes also increases predation effects on the migrating runs of juvenile fish because visual predators, such as pike, pikeperch, bass etc. are more successful at capturing prey in clear water (Gregory & Levings, 1998). The introduction of predatory species into man-made lakes has also been suggested as a reason for higher mortalities of juvenile fish in artificial lakes than natural ones (Jepsen et al., 2000).
Thermal Effects
One of the most significant results of creating lakes is a change in water temperature. Unregulated river water temperatures tend to mirror ambient air temperatures quite closely. Lakes, however, tend to reflect ambient temperatures only near the surface. Deeper lake water generally stays cool and reasonably constant throughout the year. This is mainly due to the ability of lakes to become thermally stratified in certain circumstances. Thermoclines can form at depths where wind-driven mixing of water becomes negligible. Water is densest at four degrees Celsius and consequently deep water in lakes usually stays close to this temperature. Natural lakes usually have surface outflows and, consequently, rivers that are fed by natural lakes exhibit strong seasonal variation in temperature. Rivers that are fed from hydropower reservoirs, however, are usually much cooler and have less seasonal variation in temperature. This occurs because hydropower turbine intakes are located well below the surface of the lake so that they can continue to operate when lake levels are low. Consequently, water that is drawn into the turbines and released downstream is cold and relatively unaffected by seasonal changes in ambient temperature.
Artificially cold temperatures below dams can have important implications for fish and invertebrates because they can slow the metabolism of invertebrates and fish, and inhibit growth rates (Lehmkuhl, 1972). It is also likely that invertebrates and fish use seasonal changes in temperature to cue the timing of reproduction or other seasonal behaviors. For example, Zhong & Power (1998) showed that colder than normal water temperatures delayed the spawning activity of some anadromous species in China by 20-60 days. If the natural pattern of seasonal temperatures is suppressed, the life histories of some species may become disjointed (e.g., Pardo et al., 1998). Also, surface waters of lakes can become very warm during summer months and this can have deleterious effects on returning runs of anadromous fishes that must swim through lakes (PATH, 1999; NFSC, 1999).
Greenhouse Gases
The National Hydropower Association claims that hydropower is a ‘zero air emissions’ technology that generates ‘no greenhouse gas emissions’ (NHA, 2000). This claim, however, may be quite wrong. Research in Canada has shown that the flooding of terrestrial and palustrine areas during the filling of artificial lakes can result in the production of significant quantities of carbon dioxide and methane. This occurs because of the anaerobic microbial degradation of vegetation and organic material in the flooded areas. Moreover, depending on the extent and nature of the flooded terrain, the greenhouse gas emissions (for each unit of electricity generated) for some hydropower dams are on a par with emissions produced by modern fossil fuel-based generators (Rudd et al., 1993). More recently, the World Commission on Dams (supported by the World Bank) has found that additional vegetation inputs to reservoirs supplements the greenhouse gas production as flooded vegetation is slowly depleted. Again, it was found that greenhouse gas emissions can exceed those of fossil-fuel based technologies (Fig. 1; Pearce, 2000).

Figure 1. Comparative carbon emissions from a hydroelectric dam in Brazil versus those from a coal-fired power station (from Pearce, 2000).
Water Losses
NHA also claim that hydropower does not consume the water resource. While this may be true on a large temporal and spatial scale, it is nevertheless misleading. The formation of lakes increases the surface area of river systems and this results in significantly higher evaporation rates. For example, the mean annual flow of the lower Ebro River in Spain has reduced by c. 6% due to evaporation in artificial lakes. The annual water loss from evaporation and infiltration at Lake Powell in Colorado is estimated to be c. 1.6 million acre-feet or approximately 8% of the annual flow in the Colorado River (Miller, 2000). This loss of water can have serious implications for downstream flows, particularly where other water abstractions result in very low flows, or even total dewatering of the riverbed in extreme instances. The reduction in freshwater discharge to the sea can also reduce the extent of estuarine areas with implications for species that inhabit estuaries, lagoons, and even partly enclosed seas. For example, Ibanez et al. (1998) reported that the reduced flows in the Ebro River caused the seasonal salt wedge in the Ebro Estuary to become permanent. Also, freshwater fish species in the Qiantang estuary, China, declined from 96 to 85 species following the construction of dams in the Qiantang River (Zhong and Power, 1998). However, overall production in the Qiantang Estuary increased due to an upsurge in marine fish species using the estuary (80 cf. 15 species), although this was aided by extensive artificial propagation and stocking measures. Clearly changes in freshwater and sediment regimes below dams have important impacts on community structures in downstream ecosystems.
Health Risks
Health problems may also arise where dense aquatic vegetation and large areas of still water create ideal habitats for biting insects species such as mosquitoes and biting midges, and parasitic protists such as giardia or shistosomiasis (Cogels et al., 1998). Gill parasites such as Dermocystidium, for example, can proliferate in the warm surface waters of lakes and harm fish (Joseph, 1998). The area of suitable habitat for these pests may be increased by the formation of artificial reservoirs and this may pose health risks for people and wildlife in the area.
However, marginal wetlands also provide suitable habitat for some disease vectors, such as mosquitoes, and if wetland areas become reduced by flow regulation and inundation then the lost habitat may partly counterbalance any extra health risk.
Catastrophic downstream inundation in the event of dam failure presents another issue. Such an event could be precipitated by seismic activity, aging dam materials, or extreme hydrological events. One example of this occurred on March 12, 1928, when the St. Francis Dam failed catastrophically and released a wall of water and debris (estimated to have reached 150 feet high in narrow canyons) down San Francisquito Canyon, and eventually down the Santa Clara River Valley. The flood killed an estimated 400 to 500 people. While fatal dam failures have been relatively infrequent during the past century, the potential for such failures will only increase as the number of dams increases and as dams age. Approximately 10% of dams in the USA are likely to result in the loss of life in the event of dam failure (Joseph, 1998).
Regulated Lake levels and River Flows
Normally, rivers and lakes experience a wide range of water levels (stages) depending on rainfall, snowmelt, or groundwater inputs from the catchment. During dry summer months, rainfall inputs are small and rain fed rivers will have reduced flows. Snow melt fed rivers may, however, experience relatively high flows during the early part of the summer as winter snows begin to melt. Similarly, rain-fed rivers experience higher flows during winter months and snow-melt fed rivers may have somewhat lower flows. Flood flows can periodically inundate land surrounding rivers and lakes and this can be vital for the persistence of palustrine environments such as swamps, bogs, marshes etc. Natural lakes have some ability to regulate river flows by storing some water during periods of high flow and supplementing flows during drier periods. However, this is dictated by the geography/geology of the lake and seldom approaches levels of flow regulation that occur in lakes that are managed for hydropower production.
Controlled Lake Stages and Reduced Annual/Seasonal Variation
Since the prime purpose of hydropower dams is to produce electricity, they are usually constructed so that the reservoir has a large volume. Large storage capacity is considered necessary because seasonal demand for electricity does not always coincide with periods of peak water supply. Consequently, lakes are allowed to fill during periods of high water flow and the level allowed to reduce during periods of peak electricity demand. Usually regulatory requirements place upper and lower operational lake level limits to maintain the ecological and navigational qualities of the lake, and to prevent flooding of adjacent areas. Consequently, lake levels in hydropower reservoirs vary far less than natural lakes and maximum and minimum levels are always at the same vertical position. This situation is also true for downstream river flows because large flood events are at least partially absorbed by large reservoirs.
Due to the artificially narrow range of temporarily inundated ground, large areas of marginal wetlands have dried without the semi-regular seasonal flooding (Cogels et al., 1997; Toth et al., 1997). Also, deep sediment berms have been found to form around the perimeter of artificial lakes just below the minimum lake level because water levels seldom exceed the limits allowed by regulators (Toth et al., 1997). In natural lakes this sediment is deposited across the entire width of the floodplain and serves to provide nutrients for vegetation.
Probably the most important impact of flow regulation is the loss of plant diversity around lakes and river. A large scale, long-term study of regulated and unregulated river systems in Sweden demonstrated that although plant diversity along newly regulated river systems increases in the short term, the increase is short lived and results in a long-term reduction of c 33% (Williams, 1997; Nilsson et al., 1997). Typically, species that are lost are those adapted to periodic inundation, and this parallels the reduced extent of marginal wetlands.
Downstream Flow Fluctuations
Daily variation in electricity demand mean that very large changes in the amount of water passing through hydropower turbines occur and, consequently, very large fluctuations in downstream flow rates result (e.g., Fig. 2).
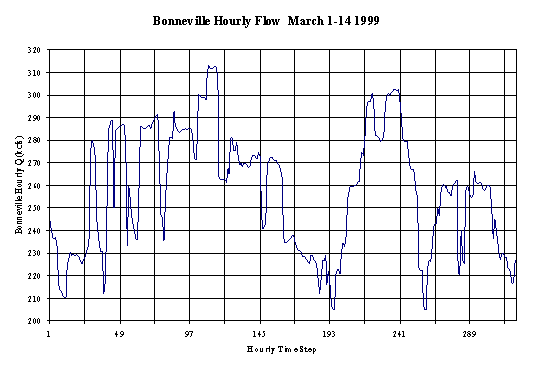
Figure 2. Downstream Flow Variation during March, 1999, below Bonneville Dam, Columbia River, Washington State, USA.
Rapidly changing flows below dams can exceed the normal rate of change found in rivers and consequently may strand fish in side channels or on gravel bars when flows are suddenly reduced. The rate of stranding increases with colder water temperatures, (Bradford, 1997) possibly because slower metabolism slows fish response. Similarly, young neotropical fishes were shown to be adversely impacted by changes in downstream flow regime. Young fish were found to be more susceptible to being swept away by increased water velocities (Ponton & Vauchel, 1998) because they could not take refuge in previously flooded off-channel habitat. This mirrors a reduction in the abundance of young fish, and in community complexity, below dams reported by Bain et al. (1988). Zhong & Power (1998) report that marked changes in the hydrological regime of the Qiantang River, China, resulted in the extinction of a highly valued fish species Macrura reevesii.
The effects of the overall reduction in downstream flows, and artificial flow patterns, are not limited to fish. Numerous studies show that macroinvertebrates are sensitive to changes in flow, and in some cases depend on seasonal flooding or low flows to cue the timing of reproduction or emergence. For example, nymphal hatching of Coloburiscoides (Ephemeroptera) appears to be delayed because of the summer release of stored water in the Mitta Mitta River in Australia (Pardo et al., 1998).
Lehmkuhl (1972) reported reduced abundance of invertebrates below dams in Saskatchewan, Canada, although this reduction was attributed to thermal effects. However, more recent research shows that invertebrate responses may differ depending on the invertebrate group. For example, lentic fauna such as bloodworms (Chironomidae) and Oxythira (Trichoptera) increase in abundance below dams as a consequence of reduced flows (and attendant changes in downstream sedimentation processes), but lotic species such as stoneflies (Plecoptera) decrease (Fjellheim et al., 1993; Mellquist, 1985). Similarly, the abundance of stonefly larvae in a Pyrenean mountain stream (France) was found to be significantly lower below a dam, and that catastrophic increases in invertebrate drift were caused by sudden increases in downstream flow (Cereghino & Lavandier, 1998). These changes reflect a shift in downstream invertebrates from lotic to lentic communities.
Miscellaneous Effects
The identification of every potential impact associated with dams is not possible in this paper since there are impacts that are not always obvious, and these may depend on the individual circumstances at a particular dam. For example, high voltage power lines carrying electricity from hydropower dams to national grids have been identified as a risk to osprey and other piscivorous raptors that hunt in rivers that have dams on them (Bird, 1983). However, this is unlikely to be an issue if these birds are not present. Likewise, navigation locks on dams near river mouths in Florida have caused the deaths of manatees (Fonferek, 1998), but this impact is only restricted to a very few dams in Florida. Clearly the individual circumstances of each dam must be considered when assessing its effects.
Summary of Dam Impacts
Rivers are highly dynamic systems that not only reflect the characteristics of their catchments (horizontal linkages) but also provide an important link between the sea and the land (longitudinal linkages). Throughout the system there are multiple ecological and physical pathways that are impacted when any one component of the system is disrupted. Figure 3 shows a hypothetical river that has been dammed and the range of potential impacts resulting from the presence of the dam. Clearly, the ecology of the river, the estuary, and even the productivity and health of the terrestrial ecosystem are intimately, and inextricably, bound together. Not one part of the ecosystem is untouched by the damming of rivers and this suggests that the removal of dams will also have very large-scale impacts. Damming rivers is not an entirely black and white issue, however, even in purely ecological terms. Dams help prevent the spread of exotic fishes and can provide new habitat for lake-loving species (including piscivorous raptors). Even so, the ecological balance tends to be more adversely impacted by the presence of dams than if they were not built at all.
The answer to the ecological problems posed by dams seems to be simple at first glance: remove them from the river. However, removing a dam is not equivalent to returning to a river to its original state. The physicochemical and biological legacies of dams can cause problems even after dams are removed, and removing dams can create new problems. This suggests that a more difficult question should be asked: which is the most ecologically favourable solution: mitigating dam effects or removing dams entirely? To answer this question, it is necessary to know the ecological costs and benefits of impacts arising from dam removals.
Move on to read about the environmental effects of removing dams